
Site menu:
主页 | 新闻 | 支原体感染症 | 医疗机构 | 相关链接 | 公司信息 |

Molecular mechanisms of lipid-antigen recognition are important in the frontier of immunology. Possible pathogeneses of autoimmune diseases and tumours now include infections with microorganisms. Therefore, the role of vaccines is increasingly important, as advancing technology has now broadened the targets of vaccination to include a greater number of infectious diseases, tumours, chronic infections, autoimmune diseases and allergies. In order to prevent infectious diseases through vaccination, it is important to identify specific antigens which often exist in the cell membrane and capsule and also become the centre of host-pathogen interactions. Structural analysis of lipid-antigens is critical for understanding the mechanisms of molecular interactions involved in the pathogenesis of immune abnormalities, and for devising strategies surrounding immune system regulation and drug discovery.
Vaccines have been used for prevention and treatment of infectious diseases for more than two centuries. Vaccination is generally considered to be the most effective method for prevention of infectious diseases. The role of vaccination has become increasingly more important due to the increasing number of targets of vaccines, including a greater number of infectious diseases, tumours, chronic infections, autoimmune diseases and allergies.
Recognition of the pathogeneses of autoimmune diseases and tumours has been changing. It is now recognized that some autoimmune diseases and tumours are caused by infections with microorganisms such as bacteria and viruses. Chronic or repetitive infections with microorganisms are triggers of chronic inflammatory diseases and the resulting appearance of downstream disorders such as autoimmune diseases and tumours. Although there are multistep processes for disease mechanisms, both environmental and genetic factors contribute to the pathogenesis of chronic inflammatory diseases, autoimmune diseases and tumours.
Vaccination, by definition, is the administration of antigenic material (the vaccine) that is designed to stimulate adaptive immunity to a particular disease. Naturally, the identification of the key antigens of microbial pathogens is an important aspect of vaccine development. An antigen is a substance (molecule) that induces production of specific antibodies (humoral immunity), activation of specific immune cells (cellular immunity) or interacts specifically with the products of the host’s immune response.
Antigenic determinants (also called epitopes), which are responsible for the specificity of the antibody, are usually parts of proteins, nucleic acids, polysaccharides, phospholipids, glycolipids or other biological macromolecules. It has become well known that different populations of T lymphocytes recognize not only peptides in the context of major histocompatability complex (MHC) class I and class II molecules, but also recognize foreign and self-lipids (lipid-antigens) in association with CD1 proteins (as CD1 proteins share structural similarities with MHC class I molecules) . Progress in the field of lipid immunology has been more recent than in the field protein immunology, and has been due largely to recent advances in lipid and membrane chemistry.
Most infectious diseases caused by bacteria begin with microbial colonization of a host surface: typically the skin, respiratory tract, gastrointestinal tract or the genitourinary tract. Humans are constantly exposed to a large number of diverse microorganisms that can cause infections. Many organisms that usually coexist harmoniously with the human host become pathogens only if the balance of the commensal relationship is disrupted. Other microorganisms are more invasive, and they overtly attack the host’s normal surface barriers and internal defence mechanisms. The human host has evolved a complex array of protective mechanisms designed to defend itself against these continuous microbial challenges.
The former paradigm was that T cells recognize peptide antigens presented by MHC class I or II molecules. Recently, it has been demonstrated that lipids can be presented by APC through the CD1 family of molecules (termed CD1-restricted antigen presentation). The structure of the CD1 family of molecules is close to that of MHC class I and II molecules (Porcelli et al., 1998). The hydrophobic tail groups of glycolipids are captured by the hydrophobic groove of CD1 molecules, similar to how hydrophobic portions of peptides are captured by the groove of MHC molecules.
Natural killer T (NKT) cells, which recognize the CD1 molecule, are a heterogeneous group of T cells that share properties of both T cells and natural killer (NK) cells. NKT differ from conventional αβ T cells in that their TCRs are far more limited in diversity and in that they recognize lipids and glycolipids presented by CD1 molecules. Like other 'unconventional' T cell subsets bearing invariant TCRs, such as CD1-restricted Natural Killer T cells, γδ T cells exhibit the more evolutionarily primitive innate immune system. The antigenic molecules that activate γδ T cells are still largely unknown. However, γδ T cells are believed to have a prominent role in recognition of lipid antigens. See also Natural Killer T Cells, Natural Killer (NK) Cells.
These lipid and glycolipid antigens include naturally occurring foreign glycolipids from intracellular pathogens and synthetic glycolipids that are related in structure to mammalian glycolipids. The portals and pockets of CD1 antigen-binding grooves influence ligand specificity and facilitate presentation of a surprisingly diverse set of antigenic lipids, glycolipids, lipopeptides and even some small, non-lipidic molecules. Presentation of antigens by CD1 proteins requires uptake and intracellular processing by APCs. There is evidence for the existence of cellular pathways that lead to presentation of both exogenous and endogenous lipid antigens
The first report on antiphospholipids was made by Wasserman in 1906 and concerned anticardiolipin antibodies in patients with syphilis. Plasma-containing proteins that prolonged phospholipid-dependent in vitro clotting assays were first described in patients with systemic lupus erythematosus (SLE) in 1952. Further addition of phospholipids to that assay system appeared to neutralize the lupus anticoagulant (LA) reaction. SLE is a disease of unknown cause in which tissues and cells are damaged by pathogenic autoantibodies and immune complexes. The malar (“butterfly”) rash is a fixed, flat or raised erythematous rash over the cheeks and bridge of the nose and can involve the chin and ears. Most SLE patients experience arthritis, glomerulonephritis and central nervous system manifestations. Anaemia is observed in most SLE patients when the disease is active.
Gangliosides, which are important constituents of the plasma membrane, are particularly abundant in the nervous system. Autoantibodies to gangliosides are found in the serum of patients with several neurological disorders such as Guillain-Barre syndrome (GBS), Fisher syndrome, sensory ataxic neuropathy and multifocal motor neuropathy. Anti-ganglioside (GD1, GM1, GQ1) and –sulfatide antibodies have been reported in these disorders. Of note, GBS and Fisher syndrome often occur after Campylobacter jejuni infections The GM1 epitope is present in LPS of the Penner 19 type, and the specific serotype Penner 19 is frequently isolated from patients with GBS. Campylobacter jejuni can be serotyped by differences in the carbohydrate structure of its LPS, which is a major constituent of the outer membrane of Gram-negative bacteria.
Guillain-Barré syndrome subsequent to Mycoplasma pneumoniae infection has also been reported to be present in GBS patients with preceding M. pneumoniae infection. Anti-Galactosylceramide (Anti-GalCer) antibody reactivity is present in GBS patients. There was a report of a patient with GBS after M. pneumoniae infection who had serum IgG antibodies to GM1b ganglioside as well as to the cold agglutinins. The cold agglutinins are polyclonal IgM autoantibodies to the I antigen of erythrocytes.
ABO blood group substances from erythrocytes were shown to be glycolipid antigens in 1965. Biochemical studies have demonstrated that many important blood group antigens are complex carbohydrates. The biosynthetic pathways that form antigens in the ABH (ABO), Lewis, P and I blood group systems are interrelated. These oligosaccharide antigens may exist free in solution. In addition, they may be covalently attached to lipid molecules (ceramide) to form glycosphingolipids. They may also be attached to polypeptides to form mucins, integral membrane glycoproteins or soluble glycoproteins.
The immune response to carbohydrate antigens, particularly when presented as repetitive epitopes, is usually independent of the thymus. In this case, multivalent antigens directly stimulate B cells to synthesize antibodies without the aid of helper T cells. Thymus-independent immune responses classically result in production of IgM antibodies. Most antibodies to carbohydrate blood group antigens are, in fact, IgM. Surprisingly, individuals lacking a carbohydrate blood group antigen on their red cells often have "naturally occurring" IgM antibodies to blood group antigens in their serum. The current understanding of this phenomenon is not that these antibodies spontaneously arise without previous antigenic stimulation. Rather, the understanding is that cross-reacting antigens are present in the environment, such as with gut bacteria, and stimulate specific IgM production.
In contrast, high-titres of IgG antibodies to carbohydrate antigens can be found in some individuals. Production of these antibodies may be stimulated by thymus-dependent forms of oligosaccharide, perhaps as individual epitopes on glycoproteins. In this way T-cells help to create a switch from production of IgM to IgG isotypes. This isotype-switching phenomenon is not yet clearly understood.
Forssman antigens have been identified from red blood cells, and are mainly responsible for haemolysis of sheep erythrocytes in the presence of globoside antiserum and complement components. The P blood group system consists of three glycosphingolipid antigens: P, P1 and pk. After microbial infection, antibodies against P antigen cause haemolysis and anaemia. The P and pk antigens have been identified as globoside and trihexosyl ceramide, respectively.
Antibodies specific for the I antigens are clinically relevant in cold-type autoimmune haemolytic anaemia. Patients with infectious diseases such as infectious mononucleosis and M. pneumoniae often develop cold agglutinins to anti-i and anti-I specificity, respectively.
Tumour-associated glycosphingolipids have attracted a great deal of interest in tumour research, particularly with regard to the potential role of tumour-associated glycosphingolipid antigens as targets for monoclonal antibodies in diagnosis and therapy of tumours. The role of glycosphingolipids has not been fully elucidated, but it has been suggested that they are involved in cellular events that may play an important biological role in tumour cell growth and in the invasive and metastatic properties of cancerous tumours (Matsuda et al., 1993).
Basically, Gram-negative bacteria contain LPS anchored by lipid A. LTA structures are present in Gram-positive bacteria, Mycoplasma, Treponema and some Gram-negative bacteriaThe lipid antigens of bacterial cell walls have adjuvant effects and specific immunogenicities that induce immune responses.





- First causative bacteria of acute bronchitis (LRTI)
- Third causative agent of pneumonia
- In every day practice the etiology of CAP is unknown in up to 70% of patients.
- Diagnosis of acute infections remains difficult.
- However, non-reliable epidemiological data because of the diagnostic problem
- To select antibiotics for lower respiratory infection (LRTI) or CAP
- To diagnose extrapulmonary manifestations, chronic and mycoplasma-related diseases
- Mycoplasma pneumoniae infection sometimes associated with serious illness.
- Because of the global spread of antimicrobial (macrolide-lincosamide) resistance is a current problem.
- Wide spreading nature because of the prolonged cough in schools, hospitals etc.
- Early diagnosis is desired also from the viewpoint of medical economics.
- a leading pathogen of both upper and lower human respiratory infections (URTI and LRTI) , in all age groups.
- known as the cause of “primary atypical pneumonia” or “community-acquired pneumonia (CAP)”.
- National Institute of Infectious Diseases (Japan)
- IDWR(Infectious Diseases Weekly Report)
- Weekly reported cases per sentinel in comparison with past 10 years
- Incidence : Each Year an estimated 1 million cases and, this year 4-5 million cases
- Center for Disease Control and Prevention (CDC) (USA)
- Home page > Disease Listing
- Each Year an estimated 2 million cases and 100,000 pneumonia-related hospitalizations occur in the United States.
- Trend : Unknown. However with improved diagnostic testing, more cases may be identified.
- Mycoplasma pneumoniae infectious diseases is not only respiratory diseases, but also diseases of the whole body.
- Mycoplasma pneumonia is frequently associated with extra pulmonary manifestations such as encephalitis.•is an exacerbating factor for asthma/COPD.
- Mycoplasma pneumonia is sometimes accompanied by other clinical syndromes: Stevens-Johnson syndrome, nephritis (including IgA nephritis), autoimmune haemolytic anaemia, meningoencephalitis, GBS and acute psychosis (Mycoplasma related diseases)
- Mycoplasma pneumoniae infection causes wide variety of disease such as asthma, neurological disorders, and arthritis. In particular, for diseases with rather chronic nature such as asthma or arthritis.
Diagnosis of Mycoplamsa infection is still not easy practically. Therefore, a reliable methodology being able to definitely exclude the infection by M. pneumoniae with higher specificity has been desired.
- For the selection of the correct antibiotics
- To see the effect of antibiotics
- To reveal the landscape of mycoplasma infection
- For the reliable epidemiological study
What performance is desired?
- As early as possible: sensitive and specific
- Able to see the clinical course: quantitative
- •Molecular detection method such as polymerase chain reaction (PCR) was expected to be a promising tool for the rapid diagnosis of M. pneumoniae infection and has been shown to be sufficiently sensitive and specific.
- However, there are some limitations for the routine use of this methodology in clinical practice; molecular detection requires DNA extraction and therefore, is not suitable for routine diagnostic laboratories, in addition to that, its sensitivity is dependent on the sample type and PCR inhibitors contained in the sample. So, even when a result of PCR is negative, it does not always mean that the patient is not infected with M. pneumoniae
- Another important point is that molecular detection cannot distinguish between infection and colonization.
- The condition of the respiratory tract sample does nor reflect the clinical state, because Mycoplasma pneumoniae infections is not only respiratory diseases, but also diseases of the whole body.
- In this respect, serology reflects the systemic reaction for M. pneumoniae infection (Beersma, 2005)(Dorigo-Zetsma, 1999). The diagnosis of M. pneumoniae infection, therefore, should be usually based on serology
- Serological methods usually include cold agglutination, complement fixation, PA, and ELISA
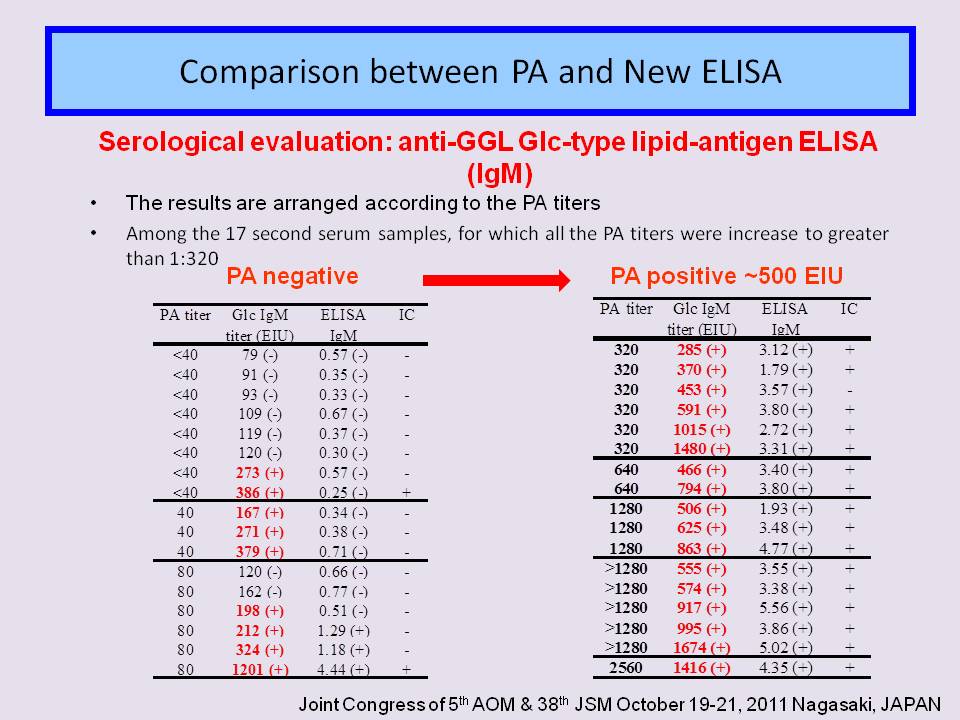
- The PA test is the most widely method used in Japan because it is easy to perform and gives semi-quantitative results with acceptable sensitivity. However, the there is some ambiguity in the visual interpretation for agglutination. The PA test is not able to distinguish between IgM and IgG class.
- To distinguish clearly current infection from past infection (clinical course), there has been a need for ELISA system to detect IgM, IgG, and IgA class antibodies separately.
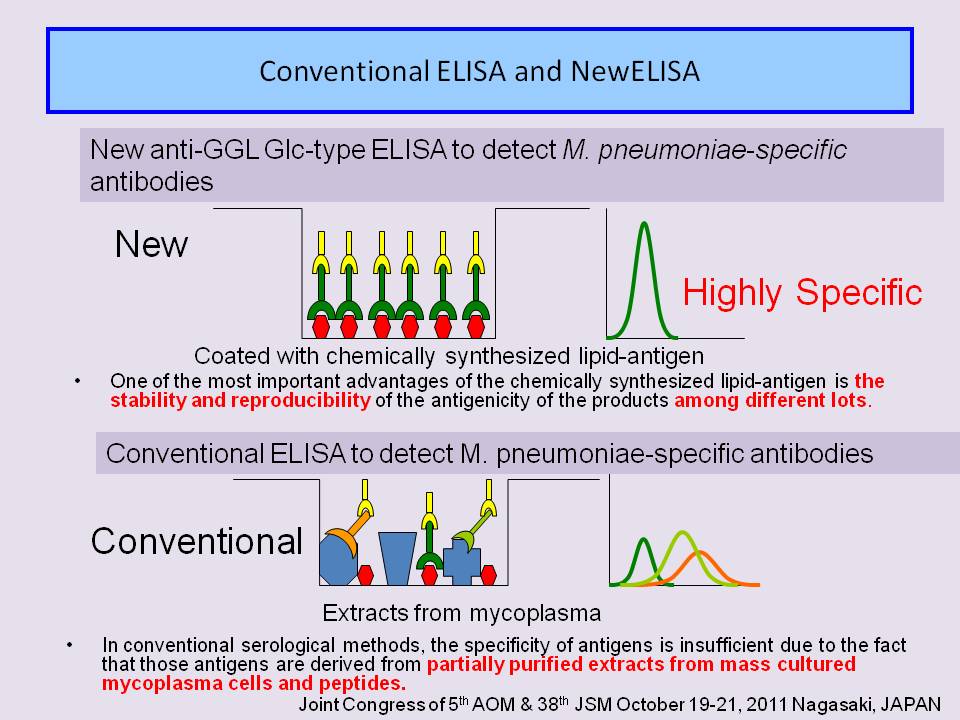
- Problem is the quality of antigens to detect anti-M. pneumoniae antibodies. However, also suffer from a problems arising from the nature of extracted antigens and non-specific reactions; the antigenicity of cellular extracts may different from each not be completely uniform in nature among lots of culture.
- The quality of Antigens is important
- These conventional methods suffer from relatively low specificity of antigens because the antigens used are derived from partially purified extracts from mass cultured mycoplasma cells.
- we focused on the small antigenic molecules of mycoplasmas, which are derived from the M. pneumoniae cell membrane component and functions as a major immunodeterminant.
What is Mycoplasma lipid-antigens?
- Cold agglutinin antibodies are found in the serum of as many as 70% of patients with mycoplasma pneumonia. The immunogenicity of M. pneumoniae is thought to be mainly due to membrane glycolipids
- The antigens of compliment fixation test (CF) are lipid antigens extracted from mycoplasma. CF is reflect the condition of adult mycoplasma infection , and related to IgG. However, t has been described that non-specific reactions, therefore, leading to the low specificity.
- Because the mycoplasma lipid-antigens have both carbohydrates as specific antigens for antibody-mediated immunity (humoral immunity) and lipid-antigen presentation (cellular immunity) specific characteristics, it is expected that these lipid-antigens could be used as therapeutic drugs.
- Microbial cell walls containing glycolipids and phospholipids are rich in antigens, which are mainly located on the surface of microbial cell membranes.
- Microbial antigens predominantly acting on antigen presentation machinery, and have a profound adjuvant effect on the immune response. This may further be linked to an intracellular trafficking pathway thought to be presented by CD1 to the specific populations of T cells.
- Microbial lipid antigens, such as LPS (endotoxin), LTAs, lipoarabinomannan (LAM) and mycoplasma lipid antigens, have activities to stimulate the immune system. These lipid antigens exert a wide variety of biological effects in humans, and are also called immunostimulants (immunomodulators).
- Too complex to utilize usually
Microbial cell walls containing glycolipids and phospholipids are rich in antigens, which are mainly located on the surface of microbial cell membranes. Microbial antigens predominantly acting on antigen presentation machinery through its ligation of Toll-like receptor, which have a profound adjuvant effect on the immune response. This may further be linked to an intracellular trafficking pathway thought to be presented by CD1 to the specific populations of T cells.
Microbial lipid antigens, such as LPS (endotoxin), LTAs, lipoarabinomannan (LAM) and mycoplasma lipid antigens, have activities to stimulate the immune system. These lipid antigens exert a wide variety of biological effects in humans, and are also called immunostimulants (immunomodulators).
Chemically synthesized natural compounds or derivatives are expected as a potential antigens and/or adjuvants for vaccine development. Synthesized Lipid A, monophosphoryl lipid A (MPL), is one of such compounds, although is not completely same as natural compounds. Conversely, the chemically synthesized mycoplasma lipid antigen is distinguishable, because it is completely same as natural compound. Since the lipid antigen is a specific antigen for both humoral immunity and cellular immunity, it is expected to be a promising strategy to develop a novel generation vaccine.
Here, it is focused on the lipid antigens of human pathogenic bacteria. Basically, Gram-negative bacteria contain LPS anchored by lipid A. LTA structures are present in Gram-positive bacteria, Mycoplasma, Treponema and some Gram-negative bacteria. The lipid antigens of bacterial cell walls have adjuvant effects and specific immunogenicities that induce immune responses.
- Mycoplasma pneumoniae lipid-antigen –GGLs : GGL Glc-type and GGL Gal-type
- M. fermentans has lipid-antigens –GGPLs: GGPL-I and GGPL-III
- Encyclopedia of Life Science
- Patents
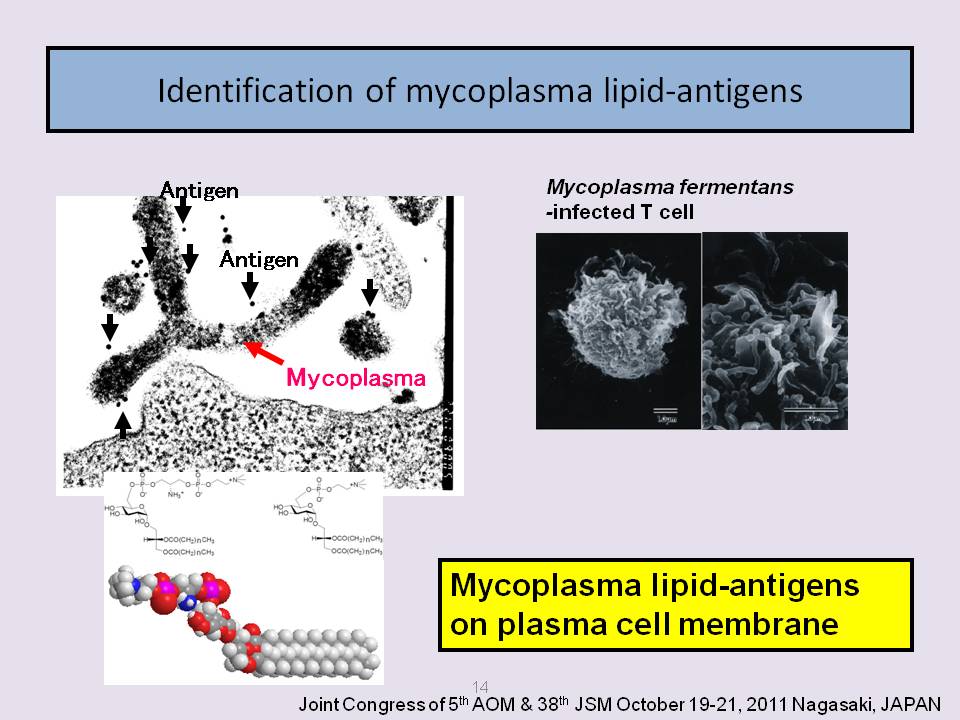
It is thought that the lipid antigens of the cell membrane of mycoplasmas play an important role in the immune response, and that the cell membrane is in direct contact with the external world. The proteins and lipids of mycoplasma membranes are the main immunogens and antigens that are recognized by anti-mycoplasma sera. Both proteins and lipids play a major role in growth and metabolic inhibition tests for mycoplasmas. Glycolipids play a main role in the assay for the M. pneumoniae and M. fermentans complement binding assays.
As a solution to the problem of a lack of antigen, small antigenic molecules of mycoplasmas have been found to be lipid-antigens. The structures of the mycoplasma lipid-antigens were then determined and chemically synthesized. M. fermentans has specific lipid-antigens called GGPL-I and GGPL-III, and M. pneumoniae also has specific lipid-antigen GGL moieties (Matsuda et al., 1994; Matsuda et al., 1997; Nishida et al., 1999; Nishida et al., 1999; Miyachi et al., 2009). The complete structures of the immunodeterminants of glycolipid antigens have been determined using instrumental analyses including nuclear magnetic resonance, mass spectrometry and chemical syntheses.
M. pneumoniae is a leading pathogen of both upper and lower human respiratory infections. Mycoplasma pneumonia, known as the cause of “primary atypical pneumonia”, is sometimes accompanied by other clinical syndromes: Stevens-Johnson syndrome, nephritis (including IgA nephritis), autoimmune haemolytic anaemia, meningoencephalitis, GBS and acute psychosis. Cold agglutinin anti-I antibodies are found in the serum of as many as 70% of patients with mycoplasma pneumonia. The immunogenicity of M. pneumoniae is thought to be mainly due to membrane glycolipids. See also Molecular Mimicry.
Mycoplasma lipid-antigens are useful biomarkers for induced levels of antibodies, and are superior in specificity and sensitivity to conventional biomarkers.
Recently, evidence has accumulated that M. fermentans is a pathogen of rheumatoid arthritis (RA). M. fermentans has frequently been isolated from the synovial fluid of patients with rheumatoid arthritis, and exhibits agglutinating activity towards erythrocytes. Moreover, M. fermentans has been shown to induce experimental arthritis in rabbits following inoculation of the trachea and knee joint (Rivera et al., 2002). GGPL, which have strong antigenicity and are species-specific immunological determinants of M. fermentans (Matsuda et al., 1997b), surprisingly have characteristic phosphocholine-containing glycoglycerolipids and can be clearly distinguished from other human related mycoplasma species through lipid analysis. Phylogenetic data strongly support the notion that mycoplasma species evolved from ancestors that are common to Gram-positive bacteria. This potential relationship suggests that it would be interesting to examine the distribution of GGPL in other microorganisms, as these lipid antigens are synthesized with specific enzymes (Fujiwara et al., 2010; Ishida et al., 2009).
- Early diagnosis of Mycoplasma pneumonie infection is important to prevent because of the appearance of antibiotics resistant mycoplasma, and also from the viewpoint of medical economics
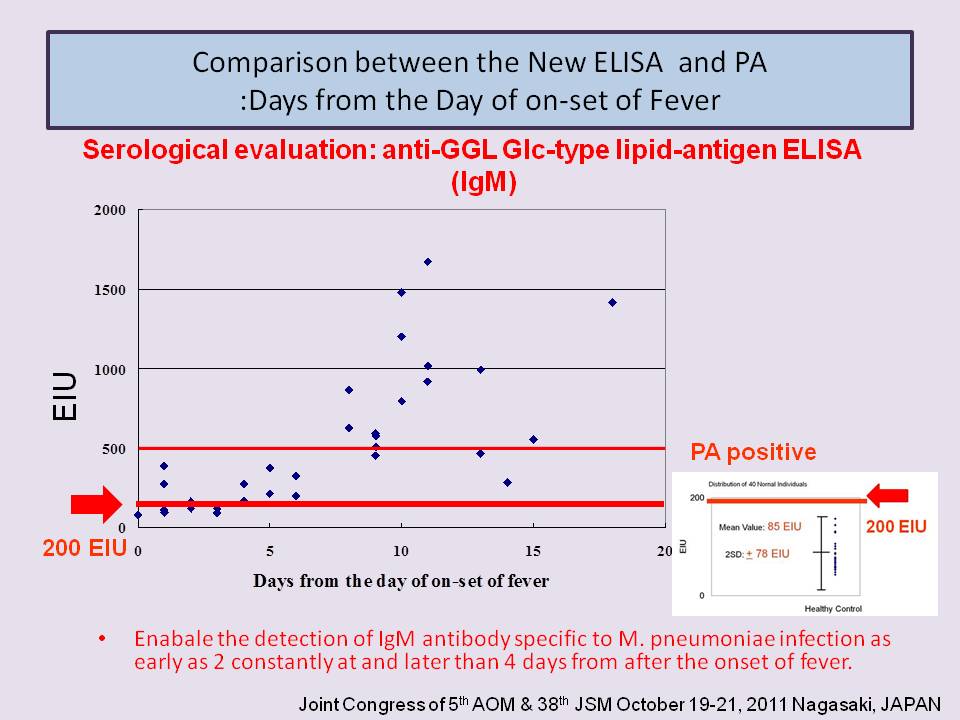
- New ELISA which utilize chemically synthesized mycoplasma species-specific lipid-antigen (anti- GGL Glc-type ELISAs ) are superior to conventional methods.
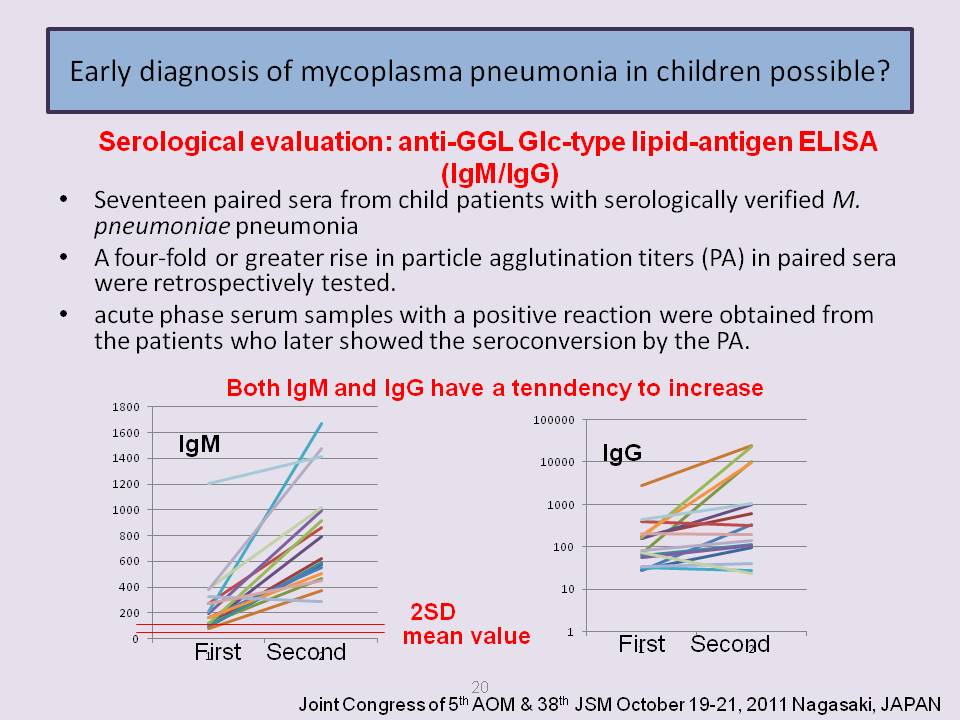
- In child patients, the anti-GGL Glc-type ELISA (IgM ) is much more suitable for the rapid early diagnosis of M pneumoniae infection than currently available conventional serological methods, and also anti-GGL Glc-type ELISA (IgG ) reflect the status of infection.
- In adult patients, the anti-GGL Glc-type ELISAs (IgM /IgG/IgA) are available to diagnose Mycoplasma pneumoniae infection suitable for the early diagnosis of M pneumoniae infection and follow the clinical course quantitatively
- The anti-GGL Glc-type ELISAs enable the studies for the links to chronic diseases, extrapulmonary diseases, and possibly M. pneumoniae infection-related disease, such as asthma rheumatoid arthritis.
Mycoplasma fermentans has been suspected as one of the causative pathogenic microorganisms of rheumatoid arthritis (RA) however, the pathogenic mechanism is still unclear. We, previously, reported that glycolipid-antigens (GGPL-I and III) are the major antigens of M. fermentans. Monoclonal antibody against the GGPL-III could detect the existence of the GGPL-III antigens in synovial tissues from RA patients. GGPL-III antigens were detected in 38.1% (32/84) of RA patient's tissues, but not in osteoarthritis (OA) and normal synovial tissues. Immunoelectron microscopy revealed that a part of GGPL-III antigens are located at endoplasmic reticulum. GGPL-III significantly induced TNF-alpha and IL-6 production from peripheral blood mononulear cells, and also proliferation of synovial fibroblasts. Further study is necessary to prove that M. fermentans is a causative microorganism of RA; however, the new mechanisms of disease pathogenesis provides hope for the development of effective and safe immunotherapeutic strategies based on the lipid-antigen, GGPL-III, in the near future. Biochem Biophys Res Commun. 2008 May 2;369(2):561-6. Epub 2008 Feb 26.
For the study of the innate immune response in infectious diseases and autoimmunity, as well as tumors, natural killer T (NKT) cells have become a major focus. Recognizing phospholipid and glycolipid antigens presented by CD1 molecules the novel T lymphocytes produce both Th1 and Th2 cytokines. Most mouse and human B cells express CD1d. Interactions between CD1d expressed on B cells and CD1d-restricted T cells may play a role in determining amount, isotype, and specificity of the antibodies produced.
Mycoplasmas have been reported as pathogens of autoimmune diseases, rheumatic diseases, asthma, neurological disorders, so on. Anti-glycolipid, such as Gal-cer, antibodies are reported to be present in the patients of autoimmune diseases with preceding mycoplasma infection. Mycoplasma lipid-antigens induce anti-mycoplasma antibodies, and they also act as autoantibodies. We have succeeded to determine the structures of the lipid-antigens, and revealed molecular mimicry. These lipid-antigens may be useful for the analysis of the signaling mechanism bridging innate and acquired immunity.
- Chemical structures, syntheses and applications of Mycoplasma pneumoniae-specific β-glycolipid antigens. 第25回国際糖質シンポジウム (2010) Matsuda, K., Ichiyama, K., Matsuda, S., Koizumi, A., Fukuda, K., Dohi, H., Harasawa, R., Saito, A., Nishida, Y.
- Mycoplasma fermentans glycolipid-antigen as a pathogen of rheumatoid arthritis. 18th Congress of The International Organization for Mycoplasmology. (2010) Kawahito, Y., Ichinose, S., Sano, H., Tsubouchi, Y., Kohno, M., Yoshikawa, T., Tokunaga, D., Hojo, T., Harasawa, R., Nakano, T., Matsuda, K.
- Novel Technologies for Mycoplasma Lipid-antigen Discovery and Vaccine Development. 18th Congress of The International Organization for Mycoplasmology. (2010) Matsuda, K.
- Novel Technologies for Mycoplasma Lipid-antigen Discovery and Vaccine Development. BIT Life Sciences’ 2nd World Congress of Vaccine. (2010) Matsuda, K.
- Development of diagnostic system based on Mycoplasma lipid-antigens. The 4th Academic Congress of Asian Organization for Mycoplasmology. (2009) Matsuda, K.
- Enzymatic synthesis of Mycoplasma fermentans specific glycoglycerophospholipid from 1,2-dipalmitoylglycerol. 109:341-345 (2010) Ishida, N., Irikura, D., Matsuda, K., Sato, S., Sono, T., Tanaka, M., Asano, K.
- Promotion of arthritis and allergy in mice by aminoglycoglycerophospholipid, a membrane antigen specific to Mycoplasma fermentans. FEMS Immunology and Medical Microbiology. 59:33-41 (2010) Sato, N., Oizumi, T., Kinbara, M., Sato, T., Funayama, H., Seiji Sato, S., Matsuda, K., Takada, H., Sugawara, S., Endo, Y.
- Variation of genes encoding GGPLs syntheses among Mycoplasma fermentans strains. J. Vet. Med. Sci. 72: 805–808 (2010) Fujiwara, M., Ishida, N., Asano, K., Matsuda, K., Nomura, N., Nishida, Y., Harasawa, R.
- Molecular cloning and expression of novel cholinephosphotransferase involved in glycoglycerophospholipid Biosynthesis of Mycoplasma fermenatans. Curr. Microbiol. 58: 535-540 (2009) Ishida, N., Irikura, D., Matsuda, K., Sato, S., Sone, T., Tanaka, M., Asano, K.
- Synthesis and absolute structures of Mycoplasma pneumoniae β-glyceroglycolipid antigens. Carbohydr. Res. 344: 36-43(2009) Miyachi, A., Miyazaki, A., Shingu, Y., Matsuda, K., Dohi, H., Nishida, Y.
- Kawahito, Y., Ichinose, S., Sano, H., Tsubouchi, Y., Kohno, M., Yoswhikawa, T., Tokunaga, D., Hojo, T., Harawsawa, R., Nakano, T., Matsuda K. Mycoplasma fermentans glycolipid-antigen as a pathogen of rheumatoid arthritis. Biochemical and Biophysical Communication 369 : 561-566 (2008)
- Matsuda, K. Lipid antigen. Encyclopedia of Life Science 2nd edition, Nature publishing group (in press)
- Matsuda, K. Phosphocholine-containing glycoglycerolipids of Mycoplasma fermentans as a pathogen of rheumatoid arthritis: Possible role of Mycoplasma fermentans GGPLs in the pathogenesis of neuroendocrine-immune abnormalities. Recent. Res. Devel. Neurosci. 1: 15-23 (2004)
- Matsuda, K., Saito, M., Yamamoto, N. Lipid antigen Encyclopedia of Life Science, Nature publishing group, vol.1 pp748-755. (2002)
- Matsuda, K., Li, J.L., Shizuko, I., Ryo, H., Saito, M., Yamamoto, N. Monoclonal antibody against Mycoplasma fermentans-specific aminoglycoglycerolipid. Microbiol. Immunol. 44: 695-702 (2000)
- Matsuda, K., Ishizuka, I., Kasama, T., Handa, S., Yamamoto, T., Taki, T. Structure of phosphocholine-containing aminoglycoglycerolipids: a novel lipid antigen of Mycoplasma fermentans. Biochim. Biophys. Acta 1349: 1-12 (1997)
- Matsuda, K., Li, J.L., Harasawa, R., Yamamoto, N. Phosphocholine-containing glycoglycerolipids (GGPL-I and GGPL-III) are species-specific major immunodeterminants of Mycoplasma fermentans. Biochem. Biophys. Res. Com. 233: 644-649 (1997)
- Matsuda, K., Harasawa, R., Li, J.L., Kasama, T., Taki, T., Handa, S., Yamamoto, N. Identification of phosphocholine-containing glycoglycerolipids purified from Mycoplasma fermentans-infected human helper T-cell culture as components of M. fermentans. Microbiolo. Immunol. 39: 307-313 (1995)
- Matsuda, K., Kasama, T., Ishizuka, I., Handa, S., Yamamoto, N., Taki, T. Structure of a novel phosphocholine-containing glycoglycerolipid from Mycoplasma fermentans. J. Biol. Chem. 269: 33123-33128 (1994)
- Matsuda, S., Matsuda, K., Ito, Y. Separation of phospholipids and glycolipids using analytical toroidal-coil countercurrent chromatography. II. Comparison of the hydrophobicity between Mycoplasma fermentans and human-brain lipids. J. Liq. Chrom. & Rel. Technol. 26: 1135-1147 (2003)
- Shingu, Y., Nishida, Y., Dohi, K., Matsuda, K., Kobayashi, K. Converient access to halide ion-catalyzed alpha-glycosylation free from noxious fumes at the donor synthesis. J. Carbohydr. Chem. 21: 605-611 (2002)
- Matsuda, K., Matsuda, S., Saito, M., Ito, Y. Separation of phospholipids and glycolipids using analytical toroidal-coil countercurrent chromatography. I. Separation of human brain lipids. J. Liq. Chrom. & Rel. Technol. 25: 1267-1281 (2002)
- Nishida, Y., Takamori, Y., Ohrui, H., Ishizuka, I., Matsuda, K., Kobayashi, K. Synthesis and absolute configulation of a novel aminoglycoglycerolipid, species-specific immunodeterminant of Mycoplasma fermentans. Tetrahedron lett. 40: 2371-2374 (1999)
- Shingu, Y., Nishida, Y., Dohi, K., Matsuda, K., Kobayashi, K. Converient access to halide ion-catalyzed alpha-glycosylation free from noxious fumes at the donor synthesis. J. Carbohydr. Chem. 21: 605-611 (2002)
- Nishida, Y., Takamori, Y., Ohrui, H., Ishizuka, I., Matsuda, K., Kobayashi, K. Synthesis and absolute configulation of a novel aminoglycoglycerolipid, species-specific immunodeterminant of Mycoplasma fermentans. Tetrahedron lett. 40: 2371-2374 (1999)
- Nishida, Y., Takamori, Y., Matsuda, K., Ohrui, H., Yamada, T., Kobayashi, K. Synthesis of artificial glycoconjugate polymer carrying 6-O-phosphocholine--D-glucopyranoside, biological active segment of main cell membrane glycolipids of Mycoplasma fermentans. J. Carbohydr. Chem. 18: 65-72 (1999)
- Li, J.L., Matsuda, K., Takagi, M., Yamamoto, N. Detection of serum antibodies against phosphocholine-containing aminoglycoglycerolipid specific to Mycoplasma fermentans in HIV-1 infected individual. J. Immunol. Methods 208: 103-113 (1997)
- Nishida, Y., Ohrui, H., Meguro, H., Ishizawa, M., Matsuda, K., Taki, T., Handa, S., Yamamoto, N. Synthesis and absolute configuration of 6-O-phosphocholine--D-glucopyranosyl glycerolipid isolated from HTLV-I-infected cell lines. Tetrahedron Lett. 35: 5465-5468 (1994)
- Matsuda, K., Taki, T., Hamanaka, S., Kasama, T., Rokukawa, C., Handa, S., Yamamoto, N. Glycosphingolipid compositions of human T-lymphotropic virus type I (HTLV-I) and human immunodeficiency virus (HIV)-infected cell lines. Biochim Biophys Acta 1168: 123-129 (1993)
- Matsuda, K., Yamamoto, N., Kaneko, T., Iwahashi, M., Hashimoto, S., Araki, K. Hypercalcemia and serum TNF-α in T-cell leukaemia. Lancet 335(8696): 1032 (1990)
- Kitamura, K., Matsuda, K., Ide, M., Tokunaga, T., Honda, M. A fluorescence sandwich ELISA for detecting soluble and cell-associated human interleukin-2. J. Immunol. Methods 121: 281-288 (1990)
- Honda, M., Kitamura, K., Matsuda, K., Yokota, Y., Yamamoto, N., Mitsuyasu, R., Chermann, J.C., Tokunaga, T. Soluble IL-2 receptor in AIDS: correlation of its serum level with the classification of HIV-induced diseases and its characterization. J. Immunol. 142: 4248-4255 (1989)